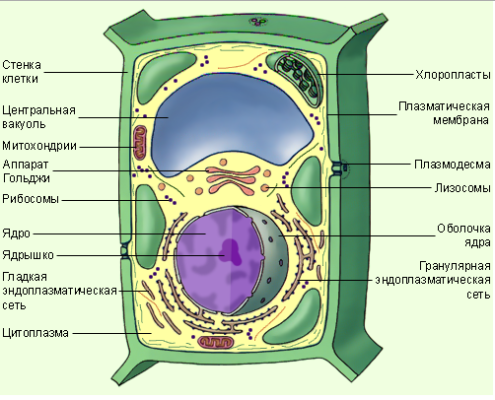
Строение клетки с подписями, Строение клеток эукариот. Цитоплазма, ядро, одномембранные органеллы

Продолжаем разговор о грибах Ликогала. Белки, которые находятся в пероксисомах, поступают туда из ЭПС и кодируются в геноме ядра. Sequential evolution of bacterial morphology by co-option of a developmental regulator.
Сферопласты патогенных бактерий называются L-формами , которые получаются в лабораториях в отсутствие клеточной стенки только в изотонических растворах [13] [14] [15]. Некоторые бактерии, паразитирующие внутри эукариотических клеток, например, микоплазма, не имеют клеточной стенки [16].
Клеточная стенка механически стабилизирована и противостоит внутреннему давлению тургорному давлению бактериальной клетки, которое составляет от 2 до 25 атм [17]. Кроме того, она играет ключевую роль в поддержании формы бактериальной клетки [18]. Через слой пептидогликана, имеющий небольшие отверстия, могут проходить только относительно небольшие молекулы массой до 50—60 кДа , причём размер проникающих молекул не зависит от толщины слоя пептидогликана. В связи с этим в тех случаях, когда через слой пептидогликана должны пройти большие молекулы, такие как белки жгутиков , пилей и ДНК при конъюгации, специфические гидролазы пептидогликана локально расширяют отверстия для их прохода [17].
В среднем нить образована 30 дисахаридами, но их количество может варьировать. К NAM присоединяется короткий пептид , состоящий из аминокислот D- глутаминовой кислоты , D- аланина и диаминопимелиновой кислоты DAP и синтезируемый вне рибосом. Аминогруппы DAP участвуют в образовании сшивок между полисахаридными цепями пептидогликана. При образовании сшивок последний D-аланин пептида высвобождается. У некоторых бактерий в пептидных мостиках присутствуют другие аминокислоты, а у грамположительных бактерий гликановые нити могут также соединяться с одним или несколькими остатками глицина в пептидных мостиках [19].
У грамположительных бактерий поверх мембраны есть от 20 до 50 нм оболочка из пептидогликана толщиной до 40 молекулярных слоёв [20]. Их положительная окраска по методу Грама связана с тем, что их толстая пептидогликановая клеточная стенка прочно связывает комплекс красителя генцианвиолета [en] с йодом , который не вымывается.
Поэтому на препаратах грамположительные бактерии выглядят фиолетовыми у грамотрицательных бактерий этот комплекс вымывается, и они приобретают цвет второго красителя, например сафранина [21].
Кроме пептидогликана, в клеточной стенке грамположительных бактерий имеются тейхоевые кислоты , которые закрепляются на поверхности клетки, образуя связи с пептидогликаном.
Липотейхоевые кислоты взаимодействуют с остатками жирных кислот клеточной мембраны. Тейхоевые и липотейхоевые кислоты представляют собой полианионы , состоящие из повторяющихся звеньев в виде фосфорилированных сахаров или остатков глицерина. Фосфатные группы в составе тейхоевых кислот могут быть заменены на глюкоуронат , в результате чего образуются тейхуроновые кислоты.
Синтез тейхуроновых кислот запускается при фосфорном голодании. Блокировка синтеза тейхоевых кислот приводит к гибели бактерий, однако конкретные функции этих соединений точно не установлены [22]. Высказываются предположения, что они действуют наподобие пружин, делая возможным растяжение и сжатие клеточной стенки. Кроме того, за счёт своей поли анионной природы тейхоевые кислоты прочно связывают ионы магния , поэтому могут выполнять в клетке роль ионообменника [23].
Поскольку у грамположительных бактерий слой пептидогликана не прикрыт сверху мембраной, перед ними стоит проблема удержания поверхностных белков. В ряде случаев поверхностные белки при помощи специальных ферментов пришиваются к фосфолипидам клеточной мембраны с образованием липопротеинов. Кроме того, поверхностные белки могут закрепляться на поверхности клетки за счёт связывания с пептидогликаном, которое обеспечивается ферментом сортазой.
Белки, предназначенные к связыванию с пептидогликаном, несут на N - и C-концах характерные последовательности, например, на N-конце находится сигнальный пептид , благодаря которому белок проходит через клеточную мембрану. Вблизи C-конца находится мотив , распознаваемый сортазой; в него вносится разрыв, после чего белок с отрезанным C-концевым фрагментом ковалентно пришивается к пептидогликану амидной связью [24].
Такую обогащённую липидами клеточную стенку иногда называют микомембраной. Микомембрана защищает бактерий от неблагоприятных условий среды и антимикробных препаратов [25]. У бактерий родов Mycobacterium , Nocardia , Corynebacterium , Rhodococcus [en] и Caseobacter в клеточной стенке обнаруживаются миколовые кислоты.
Помимо полисахаридов, у ряда патогенных грамположительных бактерий в клеточных стенках присутствуют белки, например, белок А у стафилококков, который служит важным антигеном. Кроме того, с клеточной стенкой временно, перед высвобождением в окружающую среду, связываются энтеротоксины [en] [26]. У грамотрицательных бактерий поверх клеточной мембраны тоже залегает слой пептидогликана, однако он значительно почти в 40 раз [17] тоньше, чем у грамположительных бактерий, и прикрыт сверху второй мембраной.
Клеточная и наружная мембраны отличаются по химическому составу. Пространство между клеточной и наружной мембранами называется периплазматическим пространством периплазмой [27]. В периплазматическом пространстве находится много разнообразных белков: разрушающие биологические молекулы ферменты, транспортные белки , белки, участвующие в метаболизме , а также шапероны , которые регулируют пространственную структуру других белков, защищают их от протеолиза и нежелательных взаимодействий с другими белками.
Например, в периплазме происходит образование дисульфидных мостиков и цис - транс -изомеризация пролина , которая является частью процесса созревания белка. Некоторые шапероны периплазмы участвуют в сборке ворсинок. Если под действием стрессовых условий в периплазме происходит агрегация [en] неуложенных белковых молекул, то активируется система Cpx.
Она состоит из белка CpxA, заякоренного в клеточной мембране, и связанного с ним белка CpxP, который обращён в периплазму. CpxP взаимодействует с неуложенными белками и покидает CpxA, который при этом подвергается аутофосфорилированию [en] и далее переносит фосфатную группу на цитоплазматический белок CpxR. Фосфорилированный CpxR запускает экспрессию генов стрессового ответа [28]. По мере роста клетки в периплазматическом пространстве накапливаются продукты метаболизма пептидогликана, которые клетка использует повторно [29].
Наружная мембрана состоит из двух асимметричных слоёв: внутренний слой, обращённый к клетке, состоит из фосфолипидов, а внешний — из липополисахаридов. Липополисахариды содержат О-полисахарид, коровый полисахарид и остаток липида А [en]. О-полисахарид, как правило, состоит из повторяющихся остатков галактозы , глюкозы , рамнозы и маннозы. Центральный коровый полисахарид состоит из N-ацетилглюкозамина, глюкозамина , фосфата, гептозы и кетодезоксиоктоната. Липополисахариды токсичны для животных и являются важнейшими антигенами, активирующими иммунную систему в ответ на бактериальное заражение [30].
Наружная мембрана связана со слоем пептидогликана при помощи липопротеинов, N-концы которых связаны с жирными кислотами и погружены во внешнюю мембрану, а C-концы связаны с пептидогликаном. Во внешней мембране имеются белки- порины , а также белки, связанные со сборкой поверхностных структур, конъюгацией и секрецией белковых молекул [31]. От наружной мембраны могут образовываться так называемые везикулы наружной мембраны , имеющие диаметр от 20 до нм.
В отпочковывании везикул принимает участие цитоскелет. Образование везикул может быть вызвано тем, что при росте клетки наружная мембрана увеличивается быстрее пептидогликанового слоя, а может вызываться особыми внешними условиями, например, у Porphyromonas gingivalis [en] образование везикул запускается нехваткой гемина [en].
Стенка везикул состоит из наружной мембраны, и при отпочковывании она может захватывать содержимое периплазмы. Везикулы наружной мембраны могут также служить для доставки ферментов и фрагмента наружной мембраны к клеткам-мишеням или в необходимый участок внешней среды [32]. В клетке грамотрицательной бактерии имеется от до зон слипания между наружной и клеточной мембранами, которые называют контактами Байера.
В области контактов Байера в пептидогликановом слое имеется крупное отверстие, благодаря чему наружная и клеточная мембраны могут сблизиться вплотную. Адгезию мембран могут обеспечивать компоненты некоторых экспортных комплексов. Контакты Байера могут служить для выделения наружу различных молекул, например, субъединиц пилей, кроме того, к ним прикрепляются некоторые бактериофаги [33]. Из-за наличия дополнительного барьера проницаемости наружной мембраны для достижения необходимого эффекта грамотрицательные бактерии требуют больших концентраций антибиотиков, чем грамположительные бактерии.
Наружная мембрана обеспечивает взаимодействия клеток друг с другом, с клетками организма-хозяина при патогенезе и с поверхностью субстрата. Она удерживает такие внешние структурные образования, как пили [29].
У цианобактерий поверх слоя пептидогликана располагается внешняя мембрана, однако с пептидогликаном ковалентно связаны полисахариды, из-за которых, судя по всему, цианобактерии окрашиваются по Граму положительно. Кроме того, наружная мембрана цианобактерий содержит каротиноиды [21]. Синтез пептидогликана протекает в несколько этапов. Синтез пептида начинается на NAM.
Он формируется последовательно, присоединение каждой аминокислоты катализируется определённым ферментом. Поперечные сшивки образуются с участием ферментов DD-транспептидаз [en] которые ингибируются пенициллином , которые катализируют реакцию транспептидации, сопровождающуюся высвобождением остатка D-аланина.
Образование гликановых цепей катализируют трансгликозилазы , кроме того, имеются бифункциональные ферменты, обладающие и трансгликозилазной, и транспептидазной активностями [19].
Согласно одной из предложенных моделей, при росте клетки в клеточную стенку сначала добавляются новые нити, и лишь потом происходит разрыв старых связей. У большинства бактерий в клеточной стенке находится множество ферментов автолиза , которые разрушают различные химические связи в пептидогликане [34].
Под системами секреции у бактерий понимают белковые комплексы, расположенные в клеточной мембране бактерий и служащие для выведения во внешнюю среду различных веществ.
В частности, их используют патогенные бактерии для выделения факторов вирулентности [en] преимущественно белковой природы. На основании состава, структура и действия системы секреции делят на несколько типов.
Существует по меньшей мере шесть типов систем секреции, специфичных для грамотрицательных бактерий, четыре типа систем секреции уникальны для грамположительных бактерий, а два типа систем секреции имеются у обеих групп бактерий. Типы бактериальных систем секреции и их основные свойства приведены в таблице ниже [25].
У многих бактерий поверх клеточной стенки или внешней мембраны залегает капсула, состоящая из экзополисахаридов [en].
Структурной основой капсулы служат линейные или разветвлённые полигликаны и олипептиды, состоящие из одинаковых или разных мономеров. У непатогенных бактерий капсулы служат для защиты высыхания, например, именно благодаря капсуле цианобактерии рода Nostoc могут расти в пустыне. У патогенных бактерий капсула резко увеличивает вирулентность , так как иммунная система плохо справляется с бактериями, покрытыми капсулой: они плохо связываются с антителами и не поддаются фагоцитированию [35].
Поверхность некоторых бактерий как грамположительных, так и грамотрицательных покрыта S-слоем, состоящим из упорядоченно уложенных белковых субъединиц. У бактерий очень редки случаи, когда S-слой является единственной плотной оболочкой, обычно он сосуществует вместе с пептидогликановой клеточной стенкой. S-слой не играет формообразующей роли и часто утрачивается бактериями, растущими в лабораторных условиях. Сборка S-слоя начинается с того, что его белковые субъединицы секретируются в экзоплазматический компартмент , где они спонтанно агрегируют, связываясь друг с другом гидрофобными , водородными и электростатическими связями.
S-слой обеспечивает механическую защиту бактериальной клетки, препятствует попаданию в клетку экзогенных молекул, взаимодействует с бактериофагами. У патогенных бактерий S-слой является важным фактором вирулентности [36].
Большинство бактерий подвижны, и их подвижность обеспечивается одним или нескольким жгутиками , которые представляют собой поверхностные белковые структуры. Расположение жгутиков на клетке может быть различным.
У монотрихов имеется только один жгутик, у лофотрихов на одном из полюсов клетки находится пучок жгутиков, у амфитрихов на противоположных полюсах клетки находится по одному жгутику, а у перитрихов многочисленные жгутики разбросаны по всей поверхности клетки. Длина жгутика варьирует, но диаметр обычно составляет 20 нм [37]. Основание бактериального жгутика представлено базальным телом, состоящим из двух у грамположительных или четырёх у грамотрицательных бактерий белковых колец, стержня и моторных белков.

От базального тела отходит крючок, переходящий в филамент , который завершается «шапочкой». Филамент представляет собой жёсткий цилиндр, образованный белком флагеллином. В клеточной мембране находятся кольца M и S, которые часто рассматривают как единое целое.
MS-кольцо окружено несколькими моторными белками, которые передают вращающий момент на филамент. У грамотрицательных бактерий, помимо колец M и S, есть ещё два кольца: P, залегающее в пептидогликановом слое, и L, находящееся во внешней мембране. Через все кольца проходит жёсткий стержень, передающий вращающий момент на филамент [38].
При сборке жгутика сначала в мембране клетки появляется MS-кольцо, к которому прикрепляются моторные белки, далее формируются P- и L-кольца у грамотрицательных бактерий , крючок и филамент. В такой же последовательности запускается экспрессия генов, кодирующих белки соответствующей части жгутика [39]. Через полое внутреннее пространство базального тела новые флагеллиновые субъединицы поступают к вершине растущего жгутика, где самоорганизуются по спирали.
Чтобы субъединицы флагеллина не уходили во внешнюю среду, конец растущего филамента прикрыт «шапочкой», которая не даёт им покинуть жгутик. В среднем зрелый филамент состоит из около 20 тысяч молекул флагеллина, а белки жгутика кодируются более чем 30 генами [40]. Вероятно, движущей силой вращения жгутика у бактерий является протонный градиент. Поток протонов , проходящий через кольца M и S или между базальным телом и клеточной мембраной, запускает вращение жгутика [41].

Движение клетки происходит за счёт вращения жгутика по часовой стрелке или против неё. У монотрихов клетка медленно вращается в направлении, противоположном вращению жгутика. Если жгутик вращается по часовой стрелке, то клетка движется жгутиком вперёд, а если против, то клетка выталкивается жгутиком вперёд то есть движется жгутиком назад.
Некоторые бактерии, имеющие единственный жгутик, вращают его только по часовой стрелке, и, чтобы сменить направление движения, им нужно остановиться и переориентироваться.
У перитрихов жгутики вращаются против часовой стрелки, и, если нужно сменить направление движения, клетка останавливается и совершает кувырок [41]. У некоторых бактерий рода Vibrio в частности, Vibrio parahaemolyticus [en] [42] и некоторых протеобактерий , таких как Aeromonas , имеются две различные жгутиковые системы, белковые компоненты которых кодируются различными наборами генов, а для вращения используются разные ионные градиенты. Полярные жгутики, относящиеся к первой жгутиковой системе, присутствуют постоянно и обеспечивают подвижность в потоке жидкости, а боковые жгутики, относящиеся ко второй жгутиковой системе, экспрессируются только тогда, когда сопротивление окружающей жидкости так велико, что полярные жгутики не могут вращаться.
Благодаря этому бактерии могут скользить по различным поверхностям и в вязкой жидкости [43] [44] [45] [46] [47] [48]. Пили также известны как фимбрии или ворсинки — нитевидные белковые структуры, расположенные на поверхности клеток многих бактерий. Размер пилей варьирует от долей мкм до более чем 20 мкм в длину и 2—11 нм в диаметре.
Пили участвуют в передаче генетического материала между бактериальными клетками конъюгация , прикреплении бактерий к субстрату и другим клеткам, отвечают за адаптацию организмов, служат местами прикрепления многих бактериофагов. Структурно пили могут быть от тонких нитевидных образований до толстых палочкообразных структур с осевыми отверстиями.
Пили состоят из одного или нескольких типов спирально уложенных белковых молекул, которые называют пилинами [en] или фимбринами [49].
В образовании пилей, помимо самих белков-пилинов, участвуют дополнительные белки, способствующие правильной сборке. У грамотрицательных бактерий они должны пройти через клеточную мембрану, периплазматическое пространство и наружную мембрану [50].
Иногда слизистая структура окружает не отдельную клетку, как в случае капсулы, а скопление клеток, и тогда покровную слизистую структуру называют чехлом. Чехлы могут покрывать не только вегетативные клетки, но и другие варианты дифференцированных клеток, например, покоящиеся структуры цисты, эндоспоры , гетероцисты. Чехлы имеются, например, у внутриклеточных паразитических [en] хламидий и цианобактерий [51].
У некоторых грамотрицательных бактерий, в частности, цианобактерий, псевдомонад [en] , метилотрофных бактерий, имеются особые поверхностные полые белковые структуры — шипы. Шипы представляют собой жёсткие полые конические или конусообразно-цилиндрические структуры, которые прикреплены к наружной мембране.
В среднем на клетку приходится 10 шипов. Диаметр шипов составляет 50— нм , длина — 0,5—3 мкм. Стенка шипа толщиной около 7 нм образована спирально уложенными молекулами белка спинина.
Конусообразно-цилиндрические шипы имеют открытый конец, у конических конец запечатан. Шипы образуются путём самосборки. Их функции неясны, возможно, шипы задействованы в образовании микроколоний, прикреплении к субстратам и клеткам, защите от фаготрофных протистов [52]. У бактерии Bacillus vesiculiferous , которая живёт в бедном кислородом кишечнике рачков , и некоторых других бактерий имеются так называемые газовые баллоны — крупные структуры, наполненные газом и прикреплённые к поверхности клетки.
Они представляют собой сферические, цилиндрические или игольчатые везикулы, стенка которой представлена мембраной толщиной около 2 нм. Вероятно, газовые баллоны служат для запасания кислорода [53].
У бактерий из типа Фирмикуты , способных к ферментативному разрушению целлюлозы , соответствующие ферменты — целлюлазы — находятся в особой структуре, целлюлосоме, которая находится в экзоплазматическом компартменте. Целлюлосома — очень крупный от 2 тысяч до 6,5 тысяч кДа белковый комплекс, состоящий из 14—26 белков. Целлюлосома способствует прикреплению бактерий к субстрату, обеспечивает оптимальное взаимодействие с ним целлюлаз и поступление продуктов гидролиза целлюлозы к поверхности клетки [54].
У некоторых цианобактерий в экзоплазматическом компартменте формируются кристаллы минералов , например, кальцита CaCO 3 [55]. Нуклеоид — зона неправильной формы в цитоплазме прокариотической в том числе бактериальной клетки, в которой находится геномная ДНК и ассоциированные с ней белки.
В большинстве случаев геном бактерии представлен кольцевой молекулой ДНК [en] , которую также называют хромосомой. Репликация геномной ДНК начинается от участка инициации origin of chromosomal replication , oriC , из которого две репликативные вилки движутся в противоположных направлениях и встречаются в сайте терминации Ter , в котором далее происходит размыкание сцепленных дочерних хромосом [ источник не указан дней ]. Количество хромосом в бактериальной клетке зависит не только от видовой принадлежности, но и от фазы развития популяции.
В качестве бактерий, постоянно имеющих несколько хромосом, можно привести Deinococcus radiodurans от 4 до 10 хромосом , Borrelia hermsii [en] от 8 до 16 хромосом , Desulfovibrio gigas от 9 до 17 хромосом , Azotobacter vinelandii до 80 хромосом. Молодые клетки обычно содержат больше хромосом, чем старые. Иногда хромосомы представляют не просто копии геномной ДНК: в некоторых случаях геном распределяется между несколькими неодинаковыми хромосомами и внехромосомными элементами плазмидами.
Так, у Agrobacterium tumefaciens , Brucella melitensis и Rhodobacter sphaeroides [en] имеются две разные хромосомы, а у Bacillus cereus , Leptospira interrogans [en] и Rhizobium meliloti [en] , кроме хромосом, есть одна или две мегаплазмиды, состоящие из — тысяч пар оснований п. H-NS компактизует ДНК, влияет на её сверхспирализацию, вызывает в ней изгибы, однако по большей части он задействован в экспрессии генов: под его контролем находятся до 40 генов.
Белки SMC имеются у представителей всех трёх доменов жизни. Они гомологичны миозину , у эукариот они играют роль моторов при конденсации хроматина [58]. Плазмиды — это стабильные элементы внехромосомной ДНК, которые не являются строго необходимыми для функционирования бактериальной клетки. Гены, содержащиеся на плазмидах, дают своим обладателям полезные свойства, увеличивающие их приспособленность, — например, гены, отвечающие за производство антибиотиков и устойчивость к ним; гены, повышающие устойчивость к тяжёлым металлам и УФ-излучению ; гены, кодирующие все компоненты, необходимые для конъюгации [59].
Как и у других клеток, рибосома у бактерий состоит из большой и малой субъединиц, имеющих коэффициенты седиментации 50S и 30S молекулярная масса 1,5 тысячи кДа и кДа соответственно. За исключением L7 и L12, все рибосомные белки входят в состав рибосомы и представлены одной копией. Они имеют массу от 5 до 60 кДа и состоят из 45— аминокислотных остатков [60]. Иногда гены рРНК разделены спейсерами , в состав которых могут входить гены, кодирующие сериновые , глутаматные и изолейциновые тРНК.
Долгое время считалось, что у прокариот цитоскелета нет, однако с начала х годов стали накапливаться данные о наличии у прокариот разнообразных филаментов [62]. У прокариот не только имеются аналоги ключевых белков цитоскелета эукариот, но и белки, не имеющие аналогов у эукариот [63] [64] [65] [66]. Элементы цитоскелета играют важные роли в делении клеток , защите, поддержании формы и определении полярности у различных прокариот [67] [68].
Примерами белков цитоскелета прокариот могут служить такие белки, как FtsZ [en] , гомологичный эукариотическому тубулину [69] , необходимый для цитокинеза и действующий подобно актино- миозиновому сократительному кольцу эукариот [70] ; MreB , соответствующий эукариотическому актину [71] ; кресцентин , аналогичный белкам промежуточных филаментов эукариот [9].
Большинство включений обеспечивают клетки энергией и необходимыми элементами в неблагоприятных условиях. Некоторые гранулы окружены тонкой мембраной толщиной от 2 до 4 нм ; обычно такая мембрана имеет белковую природу, но иногда содержит и липиды. Гранулы полифосфатов также называют гранулами волютина , они являются запасным источником фосфата, необходимого для синтеза нуклеотидов. Бактерии, использующие серу в своём метаболизме, содержат в цитоплазме гранулы элементарной серы, окружённые однослойной белковой мембраной.
Цианобактерии запасают цианофицин [en] , который в условиях азотного голодания служит источником этого элемента [72]. Цианофицин представляет собой полипептид с молекулярной массой от 25 до кДа , который, кроме цианобактерий, синтезируется и некоторыми другими группами бактерий, как, например, ацинетобактеры [73]. Многие бактерии содержат полиглюкозидные гранулы, состоящие из нерастворимых полиглюкозидов молекулярной массой до кДа. Полиглюкозиды бактерий представляют собой разветвлённые цепочки из остатков глюкозы, аналогичные гликогену животных.
У цианобактерии Prochloron didemni запасается аналог растительного крахмала [74]. У многих бактерий в цитоплазме содержатся полигидроксиалканоатные гранулы, которые выступают эндогенным источником углерода или энергии.
Пилигидроксиалканоаты представляют собой сложные эфиры 3-, 4- или 5- гидроксикарбоновых кислот ; они окружены белковой мембраной [75]. В редких случаях в цитоплазме бактерий обнаруживаются гидрофильные кристаллы, не окружённые мембраной. Вероятно, именно благодаря им бактерия имеет крупные размеры, так как многочисленные гранулы обеспечивают приемлемую интенсивность транспортных процессов. Некоторые анаэробные бактерии, например, Sulfospirillum barnesii , содержат кристаллы элементарного селена диаметром до нм.
Они образуются в результате восстановления ядовитого иона селенита SeO 3 У ряда бактерий, таких как Rhodopseudomonas palustris [en] и Halococcus morrhuae [en] , в цитоплазме имеются неокружённые мембраной магниточувствительные кристаллы железа в магнитосомах кристаллы железа окружены мембраной. Эти кристаллы называются параспоральными гранулами [77]. Эти органеллы обнаружены у всех цианобактерий и многих хемотрофных бактерий, фиксирующих CO 2 [78]. Карбоксисомы содержат фиксирующие углерод ферменты [79].
Они представляют собой многогранные однослойные белковые тела полиэдрической формы от 80 до нм в диаметре. Они являются основной частью механизма концентрирования CO 2 , что помогает преодолеть неэффективность рибулозодифосфаткарбоксилазы Рубиско — главного фермента, фиксирующего углерод в цикле Кальвина. Магнитосомы представляют собой окружённый мембраной цельный кристалл химически чистого магнетита Fe 3 O 4 или грейгита Fe 3 S 4.
Бактерии, обладающие магнитосомами, способны к магнитотаксису — движению, связанному с реакцией клетки на магнитное поле их называют магнитотактическими бактериями. Если говорить о строении клетки, то стоит помнить, что строение животной клетки и растительной имеют свои отличия.
Для изучения строения клетки растений ученые используют лук.

Более подробно о том, как проводится исследование, мы расскажем в другой статье. А вот изучать строение клеток животного происхождения лучше всего на кусочке мяса.
Что касается человеческих клеток, то в этом случае ученые рекомендуют использовать уже готовые препараты. На сегодня существуют такие микроскопы например Olympus BX 43 , с помощью которых удается изучить не только кровеносную и лимфатическую систему, но и клетки нервной системы, кожи, мышц и прочее.
Исследование клеток в домашних условиях можно с помощью электронного или оптического микроскопа, которые доступны каждому в любом интернет магазине. У нас Вы можете не только приобрести микроскоп, но и получить совершенно бесплатную консультацию по его выбору, узнать все характеристики интересующей Вас модели.
Для начала работы в домашних условиях идеальным решением будет микроскоп начального уровня. Но если у Вас есть возможность и опыт работы с микроскопами большого увеличения, то приобретение такого микроскопа будет не лишним. Итак, детально остановимся на изучении клетки под электронным микроскопом.
Как мы сказали уже выше, оптимальным препаратом для изучения будет клетка лука.
Поместив препарат под микроскоп обращает на себя внимание то, видны отдельные прямоугольники, между которыми определяются стенки. Это и есть не что иное, как клетка. Благодаря тому, что стенки клеток у лука плотные и упругие, они не деформируются и не изменяют свою форму.
Но есть и такие растения, у которых клеточные стенки настолько тонки и хрупкие, что легко приводит к ее повреждению. Это, например, наблюдается у апельсина. А вот клетки дуба или другого дерева разрушить намного сложнее. В каждой отдельной клетке видно содержимое, которое носит название цитоплазмы, а то пространство, что заполнено клеточным соком — это вакуоль. В центре каждой клетки видно клеточное ядро.
Если для изучения используется клетка зеленого растения, то внутри ее видны отдельные хлоропласты, принимающие участие в фотосинтезе и отвечающие за цвет растения. Клетки животного происхождения лучше всего изучать на поперечном срезе кусочка мяса. Поместив препарат под микроскоп каждый сможет увидеть клетки круглой или овальной формы, внутри которых содержаться волокна. Увидеть хлоропластов в таких клетках невозможно, так как они в них отсутствуют.
Для изучения человеческих клеток отлично подходит препарат из клеток крови. Его Вы можете найти в наборе с микроскопом, приобрести или приготовить самостоятельно.